BOLD and Brain ActivityDoes the BOLD response result from the firing of nerve cells?
|

|
Yes, although indirectly. Increased neuronal activity induces changes in regional blood flow, blood volume, and oxygen extraction through a process known as neurovascular coupling. Physiological mechanisms producing neurovascular coupling are described in the Advanced Discussion tab below.
Although the BOLD signal does relate to the number of "firing" nerve cells, it more closely correlates with extracellular local field potentials (LFPs). LFPs are slowly changing voltages recorded from large populations of brain cells with a much longer time course than depolarization "spikes". LFPs reflect the total activity of regional neural networks including not only neuronal discharges but also the sum of positive and negative post-synaptic potentials at multiple dendritic connections.
The regional BOLD response generated from a brief peripheral stimulus (such as a flashing light or pin prick) is known as the Hemodynamic Response Function (HRF). The HRF typically demonstrates a small initial dip, followed by a tall peak, and then a variable post-stimulus undershoot.
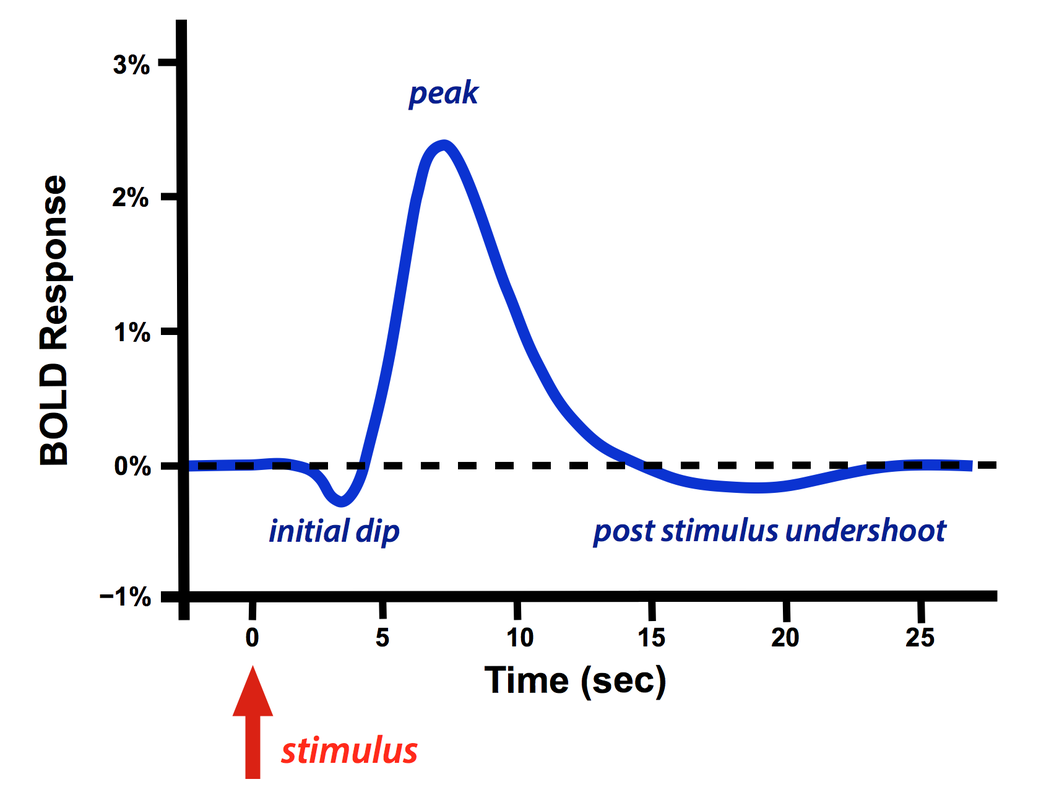 BOLD Hemodynamic Response Function (HRF)
BOLD Hemodynamic Response Function (HRF)following a single brief stimulus
The initial dip is inconsistent and variable, more commonly observed at very high fields (≥ 7.0T). Its mechanism remains disputed, with the two most popular theories being a) increased early metabolic extraction of blood oxygen, and/or b) increased local cerebral blood volume. The dip is interesting because it may offer a higher spatial specificity for activation than does the large peak.
The positive dominant peak constitutes the bulk of the BOLD response, quickly overwhelming the small initial dip. During this phase regional cerebral blood flow increases out of proportion to immediate metabolic needs. The result is that the ratio of oxyhemoglobin to deoxyhemoglobin transiently increases, with increased MR signal. Even with a very brief stimulus, the dominant HRF response is sluggish and delayed, often not occurring until 5-15 seconds later.
The positive dominant peak constitutes the bulk of the BOLD response, quickly overwhelming the small initial dip. During this phase regional cerebral blood flow increases out of proportion to immediate metabolic needs. The result is that the ratio of oxyhemoglobin to deoxyhemoglobin transiently increases, with increased MR signal. Even with a very brief stimulus, the dominant HRF response is sluggish and delayed, often not occurring until 5-15 seconds later.
The post-stimulus undershoot is variable, most commonly observed in prolonged-stimulus experiments. Its mechanism is disputed, but probably results from both slow recovery of arterial blood volume and a late stimulus- and brain-state-dependent decrease in regional cerebral blood flow.
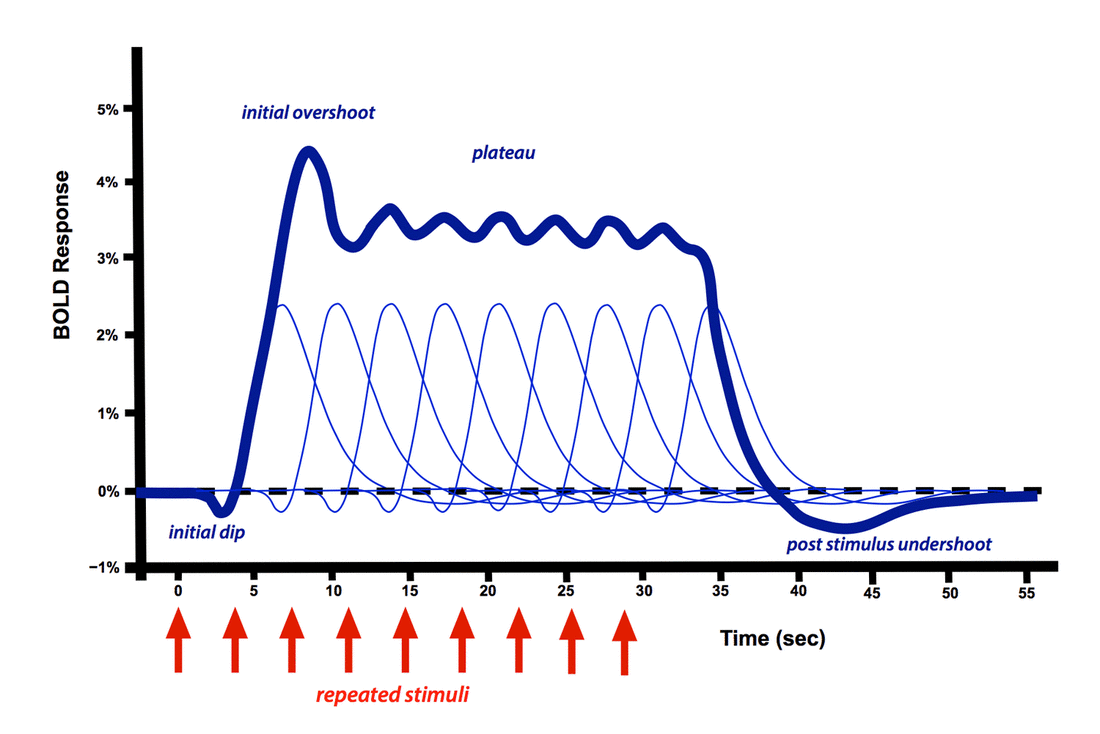
|
Multiple repeated stimuli add together in an approximately linear fashion as long as the time between stimuli exceeds 4-5 seconds. When a long train of repeated stimuli are applied, the dominant peak becomes a broad plateau, not dropping off until the stimulation ends. A small initial overshoot may sometimes be observed. Of course, considerable variation of the HRF exists between different subjects, experimental conditions, and brain regions studied.
|
|
|
More details about the HRF and BOLD response are available in the short video (left) starring Martin Lindquist from Johns Hopkins and Tor Wager from the University of Colorado at Boulder. This is part of a much larger and highly recommended video series available at their Principles of fMRI YouTube Channel accessible at this link.
|
References
Bandettini PA, Wong EC, Hinks RS, et al. Time course EPI of human brain function during task activation. Magn Reson Med 1992; 25:390–397.
Blamire AM, Ogawa S, Ugurbil K, et al. Dynamic mapping of the human visual cortex by high-speed magnetic resonance imaging. Proc Natl Acad Sci USA 1992; 89:11069–11073. (first demonstration of the single HRF from a short stimulus versus plateau-type response from much longer stimuli).
Buckner RL. Event-related fMRI and the hemodynamic response. Human Brain Mapping. 1998; 6:373–377. (shows linear response to short-term stimuli)
Chen JJ, Pike GB. Origins of the BOLD post-stimulus undershoot. Neuroimage 2009; 46:559-568.
Ekstrom A. How and when the fMRI BOLD signal relates to underlying neural activity: the danger in dissociation. Brain Res Rev 2010; 62:233-244.
Figley CR, Stroman PW. The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals. Eur J Neurosci 2011; 33:577-588. (Astrocytes outnumber of neurons by 10:1 and comprise about half of total brain volume. They play an important role in cerebrovascular regulation, neurometabolic regulation, neurotransmitter uptake, and quite possibly the BOLD fMRI signal)
Fox PT. The coupling controversy. Neuroimage 2012; 62:594-601.
Fox PT, Raichle ME. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc Natl Acad Sci USA 1986; 83:1140-1144. (Famous paper destroying the once-popular myth that CBF and metabolic rate of oxygen consumption were tightly linked).
Harris JJ, Jolivet R, Attwell D. Synaptic energy use and supply. Neuron 2012; 75:762-777. (Good review showing that over half of brain energy is used for synaptic transmission).
Hillman EMC. Coupling mechanism and significance of the BOLD signal: a status report. Annu Rev Neurosci 2014; 37:161-181.
Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. Neurophysiological investigation of the basis of the fMRI signal. Nature 2001; 412:150-157. (famous paper showing better correlation of BOLD responses with LFPs rather than action potentials)
Mullinger KJ, Mayhew SD, Bagshaw AP, Bowtell R, Francis ST. Poststimulus undershoots in cerebral blood flow and BOLD fMRI responses are modulated by poststimulus neuronal activity. Proc Natl Acad Sci USA 2013; 110:13636-41.
Phillips AA, Chan FHN, Zheng MMZ, et al. Neurovascular coupling in humans: physiology, methodological advances and clinical implications. J Cereb Blood Flow Metab 2015; Epub http://dx.doi.org/10.1177/0271678X15617954
Shmuel A, Augath M, Oeltermann A, Logothetis NK. Negative functional MRI response correlates with decreases in neuronal activity in monkey visual area V1. Nature Neurosci 2006; 9:569-577. (description of the negative BOLD response)
Sirotin YB, Das A. Anticipatory haemodynamic signals in sensory cortex not predicted by local neuronal activity. Nature 2008; 457:475-479, corrected 5 Feb 2009. (showed that vasodilation may bring additional arterial blood to cortex locally in anticipation of expected tasks)
Wager,TD, Vazquez A, Hernandez L, Noll DC. Accounting for nonlinear BOLD effects in fMRI: Parameter estimates and a model for prediction in rapid event-related studies. NeuroImage 2005; 25:206–218. (HRFs do not add linearly when events are applied at intervals less than ~2 sec)
Bandettini PA, Wong EC, Hinks RS, et al. Time course EPI of human brain function during task activation. Magn Reson Med 1992; 25:390–397.
Blamire AM, Ogawa S, Ugurbil K, et al. Dynamic mapping of the human visual cortex by high-speed magnetic resonance imaging. Proc Natl Acad Sci USA 1992; 89:11069–11073. (first demonstration of the single HRF from a short stimulus versus plateau-type response from much longer stimuli).
Buckner RL. Event-related fMRI and the hemodynamic response. Human Brain Mapping. 1998; 6:373–377. (shows linear response to short-term stimuli)
Chen JJ, Pike GB. Origins of the BOLD post-stimulus undershoot. Neuroimage 2009; 46:559-568.
Ekstrom A. How and when the fMRI BOLD signal relates to underlying neural activity: the danger in dissociation. Brain Res Rev 2010; 62:233-244.
Figley CR, Stroman PW. The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals. Eur J Neurosci 2011; 33:577-588. (Astrocytes outnumber of neurons by 10:1 and comprise about half of total brain volume. They play an important role in cerebrovascular regulation, neurometabolic regulation, neurotransmitter uptake, and quite possibly the BOLD fMRI signal)
Fox PT. The coupling controversy. Neuroimage 2012; 62:594-601.
Fox PT, Raichle ME. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc Natl Acad Sci USA 1986; 83:1140-1144. (Famous paper destroying the once-popular myth that CBF and metabolic rate of oxygen consumption were tightly linked).
Harris JJ, Jolivet R, Attwell D. Synaptic energy use and supply. Neuron 2012; 75:762-777. (Good review showing that over half of brain energy is used for synaptic transmission).
Hillman EMC. Coupling mechanism and significance of the BOLD signal: a status report. Annu Rev Neurosci 2014; 37:161-181.
Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. Neurophysiological investigation of the basis of the fMRI signal. Nature 2001; 412:150-157. (famous paper showing better correlation of BOLD responses with LFPs rather than action potentials)
Mullinger KJ, Mayhew SD, Bagshaw AP, Bowtell R, Francis ST. Poststimulus undershoots in cerebral blood flow and BOLD fMRI responses are modulated by poststimulus neuronal activity. Proc Natl Acad Sci USA 2013; 110:13636-41.
Phillips AA, Chan FHN, Zheng MMZ, et al. Neurovascular coupling in humans: physiology, methodological advances and clinical implications. J Cereb Blood Flow Metab 2015; Epub http://dx.doi.org/10.1177/0271678X15617954
Shmuel A, Augath M, Oeltermann A, Logothetis NK. Negative functional MRI response correlates with decreases in neuronal activity in monkey visual area V1. Nature Neurosci 2006; 9:569-577. (description of the negative BOLD response)
Sirotin YB, Das A. Anticipatory haemodynamic signals in sensory cortex not predicted by local neuronal activity. Nature 2008; 457:475-479, corrected 5 Feb 2009. (showed that vasodilation may bring additional arterial blood to cortex locally in anticipation of expected tasks)
Wager,TD, Vazquez A, Hernandez L, Noll DC. Accounting for nonlinear BOLD effects in fMRI: Parameter estimates and a model for prediction in rapid event-related studies. NeuroImage 2005; 25:206–218. (HRFs do not add linearly when events are applied at intervals less than ~2 sec)