Subacute Hemorrhage: Methemoglobin
Why is methemoglobin bright on T1-weighted images?
|

|

Methemoglobin (met-Hb), a brownish hemoglobin derivative, results from the oxidative denaturation of its precursor, deoxyhemoglobin (deoxy-Hb). If you have ever seen older meat change color from pink to reddish-brown you have witnessed the formation of met-hemoglobin in your very own refrigerator! The older steak (right) has a brownish hue due to the presence of met-Hb (and even more so from accumulation of its analogue, met-myoglobin.)
|
To allow reversible binding of oxygen (O2), the iron centers of deoxyhemoglobin must remain in the reduced, ferrous (Fe+2) state. Once hemoglobin has been removed from the high-oxygen environment of the circulation, however, various metabolic pathways (e.g., methemoglobin reductase) fail, allowing heme iron to become oxidized to the ferric (Fe+3) form, methemoglobin. As a result of this change in valence state, met-Hb is unable to bind oxygen. The transition from deoxy-Hb to met-Hb requires the loss of an electron from each iron center and rearrangement of electron spins in the 3d subshell. The end result is that met-Hb has 5 unpaired electrons, rendering it slightly more paramagnetic than deoxy-Hb (which has only 4 unpaired electrons). By comparison, gadolinium used for MR contrast agents has 7 unpaired electrons, the most of any element.
|

Test tube with blood sample containing 70% methemoglobin.
|
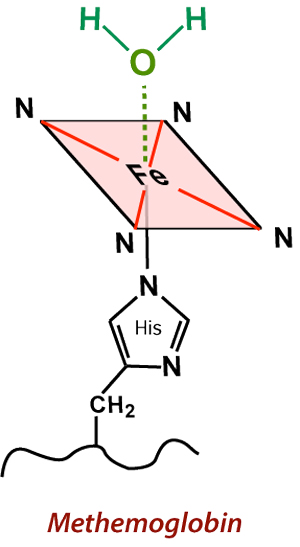
(Aquo)-methemoglobin, where H2O interacts with the Fe center, resulting in powerful inner-sphere T1-relaxation.
|
Also with the ferrous-ferric transition, the crevices in the globin subunits that previously allowed binding of oxygen are reshaped permitting access of water very close to the iron centers for the first time. The resulting complex is known as aquo-methemoglobin. This allows so-called inner sphere relaxation to occur, resulting in very short T1 values (and corresponding brightness on T1-weighted images). As long as met-Hb continues to be compartmentalized intracellularly, however, local magnetic susceptibility effects persist and the hematoma remains dark on T2/T2*-weighted images.
The subacute stage of hematoma formation is therefore distinguished chemically and radiologically by the presence and location of methemoglobin. The early subacute phase (illustrated below 2 days - 1 week) is characterized by intracellular met-Hb. The late subacute phase (1 week - 2 months, described in a later Q&A) is characterized by extracellular met-Hb.
|
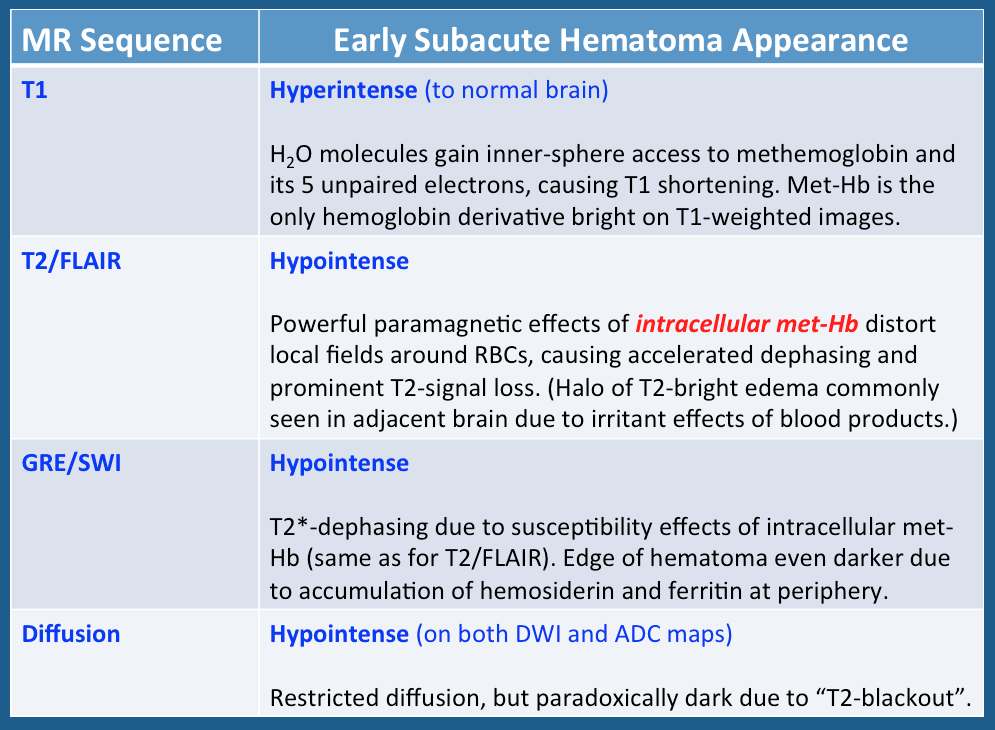
Above: Imaging characteristics of early subacute hematomas (typical time frame, 2 days - 1 week). During this stage intracellular methemoglobin predominates.
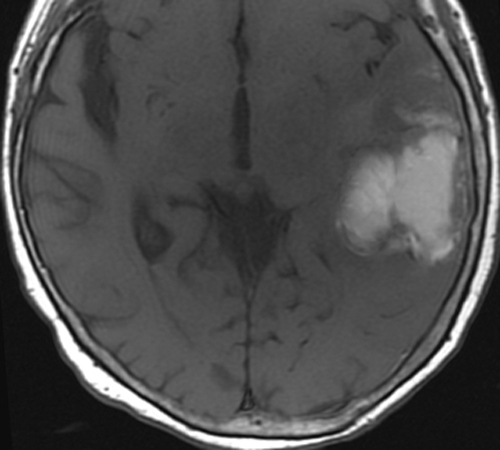
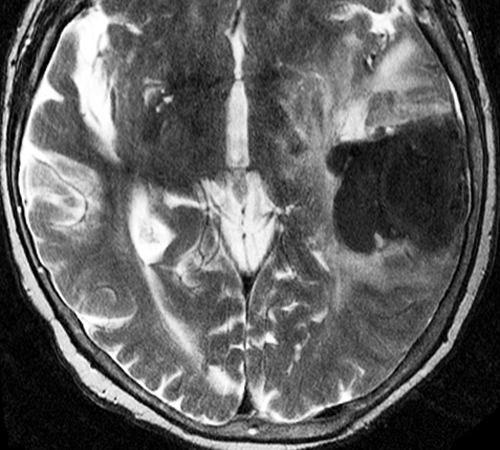
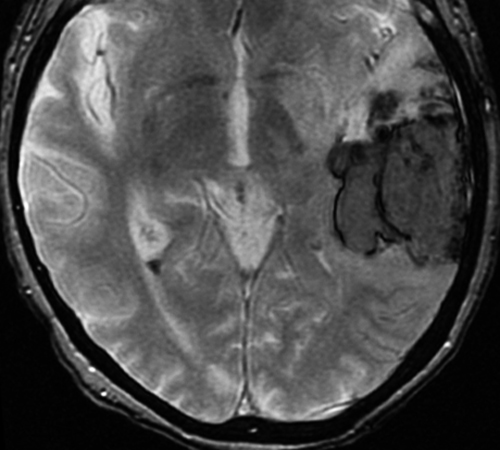
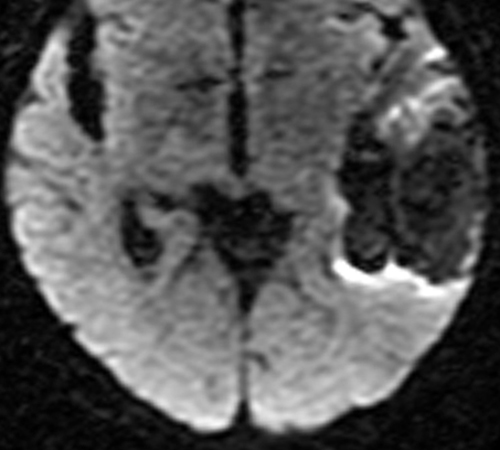
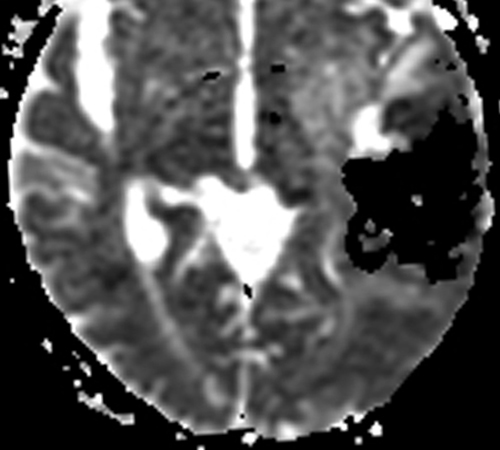
Below: MR images obtained about 3 days after hemorrhage illustrating prominent intracellular methemoglobin formation. Hematoma is hyperintense to brain on T1-weighted image (top left). Center of hematoma is markedly hypointense on T2-weighted image (top center). A halo of high signal edema in the surrounding brain is noted. SWI image (top right) shows loss of signal due to T2* dephasing from paramagnetic met-Hb confined to RBCs. Even more hypointensity is seen at periphery due to accumulation of ferritin and hemosiderin. Trace DW image (bottom left) and ADC map (bottom middle) both have dark centers, due to strong paramagnetic artifacts and the T2-blackout effect. Brighter susceptibility artifacts is present at periphery of Trace image.
Below: MR images obtained about 3 days after hemorrhage illustrating prominent intracellular methemoglobin formation. Hematoma is hyperintense to brain on T1-weighted image (top left). Center of hematoma is markedly hypointense on T2-weighted image (top center). A halo of high signal edema in the surrounding brain is noted. SWI image (top right) shows loss of signal due to T2* dephasing from paramagnetic met-Hb confined to RBCs. Even more hypointensity is seen at periphery due to accumulation of ferritin and hemosiderin. Trace DW image (bottom left) and ADC map (bottom middle) both have dark centers, due to strong paramagnetic artifacts and the T2-blackout effect. Brighter susceptibility artifacts is present at periphery of Trace image.
Important Note: The discussion above primarily applies to higher field (≥ 0.5 T) systems where imaging findings are dominated by the paramagnetic effects of hemoglobin. At low and intermediate fields some important differences in imaging appearances are noted. See this Q&A for a more detailed analysis.
References
Berman HM, Westbrook J, Feng Z, et al. The Protein Data Bank. Nucleic Acids Res 2000; 28:235-242 at www.rcsb.org (A great site well known to biochemists with over 34,000 protein sequences and 3D viewers that allow you to make images).
Chevion M, Ilan YA, Samuni A, et al. Quaternary structure of methemoglobin. Pulse radiolysis study of the binding of oxygen to the valence hybrid. J Biol Chem 1979; 254:6370-6374.
Umbreit J. Methemoglobin--it's not just blue: a concise review. Am J Hematol 2007; 82:134-144.
Yi J, Thomas LM, Richter-Addo GB. Structure of human R-state aquomethemoglobin at 2.0 Å resolution. Acta Cryst 2011; F67:1-5.
Berman HM, Westbrook J, Feng Z, et al. The Protein Data Bank. Nucleic Acids Res 2000; 28:235-242 at www.rcsb.org (A great site well known to biochemists with over 34,000 protein sequences and 3D viewers that allow you to make images).
Chevion M, Ilan YA, Samuni A, et al. Quaternary structure of methemoglobin. Pulse radiolysis study of the binding of oxygen to the valence hybrid. J Biol Chem 1979; 254:6370-6374.
Umbreit J. Methemoglobin--it's not just blue: a concise review. Am J Hematol 2007; 82:134-144.
Yi J, Thomas LM, Richter-Addo GB. Structure of human R-state aquomethemoglobin at 2.0 Å resolution. Acta Cryst 2011; F67:1-5.
Related Questions
What are the different forms of hemoglobin and why do they have different magnetic properties?
How does gadolinium cause relaxation?
What are the different forms of hemoglobin and why do they have different magnetic properties?
How does gadolinium cause relaxation?